

When the word hermaphrodite
is uttered, it is often in hushed tones with references to grotesque,
disparate rumors. In the world of biology, however, hermaphroditism is known to
be a common and successful reproductive strategy among a wide variety of
organisms. Plants, invertebrates, and many species of fish use hermaphroditism
to ensure that their genes make it on to the next generation of individuals.
This term in itself is not easily defined as there are many different forms;
sequential, simultaneous, and self-fertilizing are a few of many broad
classifications. In general, a hermaphrodite is an organism with both male and
female sex organs during some stage in its life cycle. In order to fully
understand hermaphroditism in fish and its evolutionary advantages, or lack
thereof, we must look at the different forms and the varying ways that fishes
use them. Hermaphroditism in fishes comes at a price for those that use it, but
for many species it is the only way to ensure survival in a harsh world.
 |
|
A
fantastical depiction of a hermaphrodite in Le Louvre. (Photo by Paul H.)
|
Most common in fishes is sequential hermaphroditism,
meaning that the fish starts life as one gender and at some point, due to
genetic or environmental factors, morphs into the opposite sex. The process of
changing sex from female to male is called protogyny, and this is the most
widely used form of sequential hermaphroditism (75%). The other form, switching
from male to female, is termed protandry (25%). Groupers (Serranidae), porgies
(Sparidae), wrasses (Labridae), parrotfishes (Scaridae), angelfishes
(Pomacanthidae), and gobies (Gobiidae) are all fish that are protogynous. For
example, moon wrasse populations are made up of drab females, drab primary
males, and gaudy secondary protogynous males. A secondary male, which began
life as a female, may control a harem of females and mate with them individually
while the primary males, each born with a single set of male reproductive
organs, must aggregate in large groups to spawn with a single female (Robertson
and Choat 1974). Examples of protandrous fishes are damselfishes
(Pomacentridae). Several species of clownfish live in small hierarchical groups
in a single anemone. There is a dominant female, a smaller mating male, and several
non-reproducing males with no functional gonads. Only when one of the mating
pair dies does the next highest ranking male step up and take on the sexual
transformation (Fricke and Fricke 1977). Sequential hermaphroditism is an
advantage for many fishes because it allows them to overcome the challenges of
population structure biases.
 |
|
Clownfish
are protandrous simultaneous hermaphrodites. (Image from
www.breedclownfish.com)
|
Simultaneous hermaphroditism is exactly as it sounds:
the fish has both male and female gonads at the same time. While this is
relatively uncommon in fishes, there are several species that exhibit this
characteristic in the family Serranidae. These fish may take turns fulfilling
each role over the course of multiple mating events. For these groups of fishes,
this life history could be the only way to ensure the survival of a population.
Simultaneous hermaphroditism is an incredibly valuable trait when mating
opportunities are rare due to sparse or dispersed individuals—it ensures that
when two fish do cross paths, they are always compatible. It also balances the
cost of paternal and maternal energy allocation when resources are in short
supply. These fish can be said to be the ultimate compromisers and
opportunists. While there are obvious benefits for some species of fish in
extreme situations, simultaneous hermaphroditism can be difficult to develop
because individuals must evolve congruent yet identical genitalia (Michiels
1998).
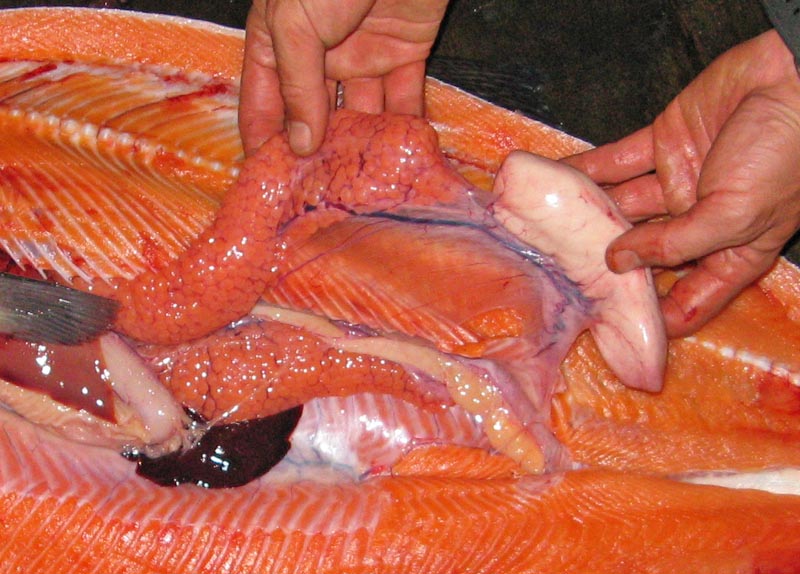 |
| Internal anatomy of a simultaneously hermaphroditic salmon. (Image from http://www.piscatorialpursuits.com |
It is rare to find a fish in nature that is a
self-fertilizing simultaneous hermaphrodite. When an organism uses its own
sperm to fertilize its own eggs, this process is called selfing and results in
genetically identical offspring, also known as clones. Far from science
fiction, this act insures that an individual’s genes are passed on regardless
of circumstance. The Mangrove Killifish, for example, is a self-fertilizing
hermaphrodite, but there are still some male-only fish in the population. While
the mangrove killifish are able to survive and reproduce on their own no matter
the conditions, it is the breeding with the males that introduces genetic
diversity and ensure the survival and adaptability of the population as a whole
(Grageda, et al. 2005). Without this outcrossing
of genes, the Mangrove Killifish would vulnerable to extinction due to a very
small gene pool. As we can see here, cloning is a useful strategy in situations
where fish are far and few between, but it isn’t a proper substitute for traditional
breeding practices.
 |
| Self-fertilizing Mangrove Killifish Rivulus marmoratus Photo http://www.practicalfishkeeping.co.uk |
In conclusion, hermaphroditism in its many forms is a
unique adaptation evolved over the course of history to increase the
survivorship of the species that use these methods. In populations of fish
where individuals are less likely to come in contact with one another,
simultaneous hermaphroditism raises the probability of an encounter with
another fish being breeding compatible from 50% all the way to 100%. In
sequential species, hermaphroditism plays a crucial role in behavioral rituals
and sex ratios. Most oddly of all, self-fertilizing hermaphrodites are able to
create a prodigy of their own even if they never come in contact with another
of their species throughout their entire lives. Many of these strategies come
with trade-offs of their own, especially the genetic issues with selfing
hermaphroditism, but in the end it is better to have a compromised existence
than none at all. Natural selection has given these species of fish what they
need to endure in even the most desperate of circumstances.
References
Fricke,
H., and S. Fricke. 1977. Monogamy and sex change by aggressive dominance in
coral reef fish. Nature 266:830-832.
Grageda, V. C., et al. 2005. Differences in life-history traits in two clonal strains of the self-fertilizing fish, Rivulus marmoratus. Environmental Biology of Fishes 73:427-436.
Michiels,
N. C. 1998. Mating conflicts and sperm competition in simultaneous
hermaphrodites. Pages 219-254 in T.
R. Birkhead and A. P. Moller, editors. Sperm competition and sexual selection.
Academic Press, San Diego, California.
Robertson,
D. R., and J. H. Choat. 1974. Protogynous hermaphroditism and social systems in
labrid fish. Proceedings of the Second International Coral Reef Symposium
1:217-225.
The diet that you eat has a huge impact on your libido and fertility. There’s no need to make major changes and turn your nutritional habits around. Still, getting rid of a few bad foods and adding certain healthy meals can make a difference.
ReplyDeleteIf you want to see incredible sexual enhancement from your routine, diet should be one of the first things to focus on. The following guide will acquaint you with the most important rules and list the foods that will speed up the outcome of your program. full prosolution review here
Size increase pills work by increasing the flow of the blood to the penis. When more blood enters the corpora cavernosa, the penis looks bigger and the erection lasts for a longer period of time.
ReplyDeletedefinite natural gain pus website